hal_131977 Co-expression
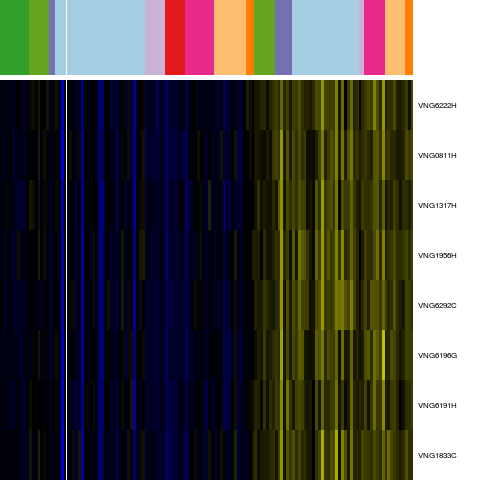
Genes in hal_131977
| Organism | Gene name | Common name | Accession | Description | Start | Stop | Strand | Chromosome |
|---|---|---|---|---|---|---|---|---|
| Halobacterium salinarum sp. NRC-1 | VNG0811H | VNG0811H | NP_279789.1 | Vng0811h | 609288 | 608896 | - | NC_002607.1 |
| Halobacterium salinarum sp. NRC-1 | VNG1317H | VNG1317H | NP_280179.1 | Vng1317h | 984111 | 983914 | - | NC_002607.1 |
| Halobacterium salinarum sp. NRC-1 | VNG1833C | VNG1833C | NP_280561.1 | Vng1833c | 1353444 | 1352827 | - | NC_002607.1 |
| Halobacterium salinarum sp. NRC-1 | VNG1956H | VNG1956H | NP_280657.1 | Vng1956h | 1443507 | 1444004 | + | NC_002607.1 |
| Halobacterium salinarum sp. NRC-1 | VNG6191H | VNG6191H | NP_395722.1 | Vng6191h | 161740 | 161471 | - | NC_002608.1 |
| Halobacterium salinarum sp. NRC-1 | VNG6196G | phoT2 | NP_395726.1 | sodium-dependent phosphate transporter | 162665 | 163411 | + | NC_002608.1 |
| Halobacterium salinarum sp. NRC-1 | VNG6222H | VNG6222H | NP_395746.1 | Vng6222h | 183053 | 183622 | + | NC_002608.1 |
| Halobacterium salinarum sp. NRC-1 | VNG6292C | VNG6292C | NP_395794.1 | Vng6292c | 223370 | 222849 | - | NC_002608.1 |
Conditions in hal_131977
| Condition ID | Condition Name |
|---|---|
| hal_101 | circadian_drk7_cycling_1980min_vs_NRC-1d.sig |
| hal_102 | circadian_drk7_cycling_2160min_vs_NRC-1d.sig |
| hal_1051 | O2_0258_-0001_L2H_vs_NRC1d.sig |
| hal_1052 | O2_0258_0005_H_vs_NRC1d.sig |
| hal_1053 | O2_0258_0010_H_vs_NRC1d.sig |
| hal_1055 | O2_0258_0030_H_vs_NRC1d.sig |
| hal_1056 | O2_0258_0045_H_vs_NRC1d.sig |
| hal_1058 | O2_0258_0180_H_vs_NRC1d.sig |
| hal_107 | circadian_drk7_cycling_3060min_vs_NRC-1d.sig |
| hal_108 | circadian_drk7_cycling_3240min_vs_NRC-1d.sig |
| hal_1138 | PQ_Const._set_1_0.25mM_-001m_vs_NRC-1f.sig |
| hal_1139 | PQ_Const._set_1_0.25mM_000m_vs_NRC-1f.sig |
| hal_1141 | PQ_Const._set_1_0.25mM_010m_vs_NRC-1f.sig |
| hal_1142 | PQ_Const._set_1_0.25mM_020m_vs_NRC-1f.sig |
| hal_1143 | PQ_Const._set_1_0.25mM_040m_vs_NRC-1f.sig |
| hal_1144 | PQ_Const._set_1_0.25mM_080m_vs_NRC-1f.sig |
| hal_1145 | PQ_Const._set_1_0.25mM_160m_vs_NRC-1f.sig |
| hal_1165 | 20060818_KO_ura3_rep2_0.189_vs_NRC-1e.sig |
| hal_1167 | 20060818_KO_ura3_rep2_0.828_vs_NRC-1e.sig |
| hal_1205 | 20060717_Cmyc_tbpF_rep1_0.401_vs_NRC-1d.sig |
| hal_1206 | 20060717_Cmyc_tbpF_rep1_0.829_vs_NRC-1d.sig |
| hal_127 | circadian_20051021_cycling_dark_42oC_1440min_vs_NRC-1d.sig |
| hal_1276 | 20060308_KO_tbpA_rep1_0.398_vs_NRC-1e.sig |
| hal_1277 | 20060308_KO_tbpA_rep1_0.813_vs_NRC-1e.sig |
| hal_128 | circadian_20051021_cycling_dark_42oC_1615min_vs_NRC-1d.sig |
| hal_129 | circadian_20051021_cycling_dark_42oC_1800min_vs_NRC-1d.sig |
| hal_131 | circadian_20051021_cycling_dark_42oC_2160min_vs_NRC-1d.sig |
| hal_1324 | 20050620_Cmyc_tfbG_rep2_0.170_vs_NRC-1d.sig |
| hal_1326 | 20050620_Cmyc_tfbG_rep2_1.000_vs_NRC-1d.sig |
| hal_133 | circadian_20051021_cycling_dark_42oC_2515min_vs_NRC-1d.sig |
| hal_1339 | 20050620_Cmyc_tfbD_rep1_0.156_vs_NRC-1d.sig |
| hal_134 | circadian_20051021_cycling_dark_42oC_2705min_vs_NRC-1d.sig |
| hal_1343 | 20050620_Cmyc_rep2_0.457_vs_NRC-1d.sig |
| hal_1344 | 20050620_Cmyc_rep2_1.072_vs_NRC-1d.sig |
| hal_135 | circadian_20051021_cycling_dark_42oC_2880min_vs_NRC-1d.sig |
| hal_1377 | NRC-1.2_0.1684_vs_NRC-1_h1.sig |
| hal_1378 | NRC-1.2_0.3063_vs_NRC-1_h1.sig |
| hal_1398 | NRC-1_0.3279_vs_NRC-1_h1.sig |
| hal_1428 | sDura3D1179_pCu_d0.100mM_t+060m_vs_NRC-1h1.sig |
| hal_1429 | sDura3D1179_pCu_d0.100mM_t+180m_vs_NRC-1h1.sig |
| hal_1453 | sDura3D1179_pZn_d0.020mM_t+060m_vs_NRC-1h1.sig |
| hal_1456 | sDura3D1179_pZn_d0.050mM_t+015m_vs_NRC-1h1.sig |
| hal_1457 | sDura3D1179_pZn_d0.050mM_t+030m_vs_NRC-1h1.sig |
| hal_1458 | sDura3D1179_pZn_d0.050mM_t+060m_vs_NRC-1h1.sig |
| hal_1459 | sDura3D1179_pZn_d0.050mM_t+180m_vs_NRC-1h1.sig |
| hal_1460 | sDura3D1179_pZn_d0.050mM_t-015m_vs_NRC-1h1.sig |
| hal_148 | circadian_dark3_ctrl_1380min_vs_NRC-1d.sig |
| hal_1481 | sDura3_pZn_d0.005mM_t+015m_vs_NRC-1h1.sig |
| hal_1483 | sDura3_pZn_d0.005mM_t+060m_vs_NRC-1h1.sig |
| hal_1484 | sDura3_pZn_d0.005mM_t+180m_vs_NRC-1h1.sig |
| hal_1485 | sDura3_pZn_d0.005mM_t-015m_vs_NRC-1h1.sig |
| hal_1491 | sDura3_pZn_d0.050mM_t+015m_vs_NRC-1h1.sig |
| hal_1492 | sDura3_pZn_d0.050mM_t+030m_vs_NRC-1h1.sig |
| hal_1493 | sDura3_pZn_d0.050mM_t+060m_vs_NRC-1h1.sig |
| hal_1494 | sDura3_pZn_d0.050mM_t+180m_vs_NRC-1h1.sig |
| hal_150 | circadian_dark3_ctrl_1500min_vs_NRC-1d.sig |
| hal_151 | circadian_dark3_ctrl_1620min_vs_NRC-1d.sig |
| hal_152 | circadian_dark3_ctrl_1800min_vs_NRC-1d.sig |
| hal_154 | circadian_dark3_ctrl_2160min_vs_NRC-1d.sig |
| hal_156 | circadian_dark3_ctrl_2340min_vs_NRC-1d.sig |
| hal_158 | circadian_dark3_ctrl_2700min_vs_NRC-1d.sig |
| hal_159 | circadian_dark3_ctrl_2880min_vs_NRC-1d.sig |
| hal_160 | circadian_drk8_control_0000min_vs_NRC-1e.sig |
| hal_162 | circadian_drk8_control_0360min_vs_NRC-1e.sig |
| hal_163 | circadian_drk8_control_0540min_vs_NRC-1e.sig |
| hal_164 | circadian_drk8_control_0720min_vs_NRC-1e.sig |
| hal_165 | circadian_drk8_control_0900min_vs_NRC-1e.sig |
| hal_166 | circadian_drk8_control_1080min_vs_NRC-1e.sig |
| hal_167 | circadian_drk8_control_1260min_vs_NRC-1e.sig |
| hal_169 | circadian_drk8_control_1620min_vs_NRC-1e.sig |
| hal_170 | circadian_drk8_control_1800min_vs_NRC-1e.sig |
| hal_171 | circadian_drk8_control_1980min_vs_NRC-1e.sig |
| hal_172 | circadian_drk8_control_2160min_vs_NRC-1e.sig |
| hal_174 | circadian_drk8_control_2520min_vs_NRC-1e.sig |
| hal_175 | circadian_drk8_control_2700min_vs_NRC-1e.sig |
| hal_178 | circadian_drk8_control_3240min_vs_NRC-1e.sig |
| hal_230 | Circadian_Dark10_Cycling_-2873min_vs_NRC-1h1.sig |
| hal_232 | Circadian_Dark10_Cycling_-2490min_vs_NRC-1h1.sig |
| hal_238 | Circadian_Dark10_Cycling_-1410min_vs_NRC-1h1.sig |
| hal_239 | Circadian_Dark10_Cycling_-1245min_vs_NRC-1h1.sig |
| hal_242 | Circadian_Dark10_Cycling_-0690min_vs_NRC-1h1.sig |
| hal_243 | Circadian_Dark10_Cycling_-0525min_vs_NRC-1h1.sig |
| hal_245 | Circadian_Dark10_Cycling_-0165min_vs_NRC-1h1.sig |
| hal_246 | Circadian_Dark10_Cycling_0000min_vs_NRC-1h1.sig |
| hal_249 | Circadian_Dark10_Cycling_0560min_vs_NRC-1h1.sig |
| hal_251 | Circadian_Dark10_Cycling_0915min_vs_NRC-1h1.sig |
| hal_252 | Circadian_Dark10_Cycling_1100min_vs_NRC-1h1.sig |
| hal_253 | Circadian_Dark10_Cycling_1270min_vs_NRC-1h1.sig |
| hal_259 | Circadian_Dark10_Cycling_2345min_vs_NRC-1h1.sig |
| hal_262 | Circadian_Dark10_Cycling_2895min_vs_NRC-1h1.sig |
| hal_263 | Circadian_Dark10_Cycling_3070min_vs_NRC-1h1.sig |
| hal_264 | Circadian_Dark10_Cycling_3252min_vs_NRC-1h1.sig |
| hal_266 | Circadian_Dark10_Cycling_3630min_vs_NRC-1h1.sig |
| hal_270 | Circadian_Dark10_Cycling_4330min_vs_NRC-1h1.sig |
| hal_285 | Cu_020_vs_NRC-1.sig |
| hal_288 | Cu_160_vs_NRC-1.sig |
| hal_363 | Cu_ura3_KO_set2_010min_vs_NRC-1h1.sig |
| hal_366 | Cu_ura3_KO_set2_080min_vs_NRC-1h1.sig |
| hal_367 | Cu_ura3_KO_set2_160min_vs_NRC-1h1.sig |
| hal_369 | Cu_vng700_KO_set2_000min_vs_NRC-1h1.sig |
| hal_371 | Cu_vng700_KO_set2_010min_vs_NRC-1h1.sig |
| hal_375 | Cu_vng700_KO_set2_160min_vs_NRC-1h1.sig |
| hal_462 | feso4__-0001m_vs_NRC-1 |
| hal_463 | feso4__0000m_vs_NRC-1 |
| hal_465 | feso4__0010m_vs_NRC-1b |
| hal_468 | feso4__0025m_vs_NRC-1b |
| hal_469 | feso4__0040m_vs_NRC-1 |
| hal_470 | feso4__0080m_vs_NRC-1 |
| hal_471 | feso4__0160m_vs_NRC-1 |
| hal_472 | feso4__0320m_vs_NRC-1 |
| hal_486 | idr1_60min_+Fe_vs_NRC1-h1.sig |
| hal_512 | tfbG_CHIS_over |
| hal_513 | tfb_D_vs_NRC-1c |
| hal_514 | tfb_C_vs_NRC-1c |
| hal_515 | tfb_B_vs_NRC-1c |
| hal_516 | tfb_A_vs_NRC-1c |
| hal_551 | NRC-1__HO_D_vs_NRC-1 |
| hal_554 | NRC-1__LO_L_vs_NRC-1 |
| hal_584 | gamma4__0000gy-010min |
| hal_585 | gamma4__0000gy-020min |
| hal_586 | gamma4__0000gy-030min |
| hal_587 | gamma4__0000gy-040min |
| hal_588 | gamma4__0000gy-050min |
| hal_589 | gamma4__0000gy-060min |
| hal_591 | gamma4__0000gy-240min |
| hal_729 | PQ_Recov._set_2_0mM_020min_vs_NRC-1f.sig |
| hal_750 | H2O2_Const._set_1_0mM_000min_vs_NRC-1f.sig |
| hal_751 | H2O2_Const._set_1_0mM_030min_vs_NRC-1f.sig |
| hal_833 | NRC-1_control_2days_vs_NRC-1h1.sig |
| hal_919 | PQ_Recov._set_2_4mM_000min_vs_NRC-1f.sig |
| hal_920 | PQ_Recov._set_2_4mM_010min_vs_NRC-1f.sig |
| hal_924 | PQ_Recov._set_2_4mM_050min_vs_NRC-1f.sig |
| hal_926 | PQ_Recov._set_2_4mM_120min_vs_NRC-1f.sig |
| hal_927 | PQ_Recov._set_2_4mM_240min_vs_NRC-1f.sig |
| hal_949 | PQ_Recov._set_1_4mM_020min_vs_NRC-1f.sig |
| hal_95 | circadian_drk7_cycling_0900min_vs_NRC-1d.sig |
| hal_950 | PQ_Recov._set_1_4mM_030min_vs_NRC-1f.sig |
| hal_969 | KO_trh6.1_0.1689_vs_NRC-1_h1.sig |
| hal_97 | circadian_drk7_cycling_1260min_vs_NRC-1d.sig |
| hal_970 | KO_trh6.1_0.3532_vs_NRC-1_h1.sig |
| hal_971 | KO_trh6.1_0.4762_vs_NRC-1_h1.sig |
| hal_972 | KO_trh6.1_0.7964_vs_NRC-1_h1.sig |
| hal_99 | circadian_drk7_cycling_1620min_vs_NRC-1d.sig |
De novo detected cis-regulatory motifs
| CRM ID | eval | PSSM |
|---|---|---|
| hal_131977_1 | 230.0000 |  |
| hal_131977_2 | 2500.0000 |  |
GREs assigned to hal_131977
| GRE ID | PSSM |
|---|---|
| hal_0 |  |
| hal_0 | |
Corems associated with hal_131977
| Corem |
|---|
| hc15251 |
| hc16269 |
| hc18595 |
| hc18596 |
| hc23879 |
| hc27530 |
| hc35295 |
| hc35473 |
| hc36106 |
| hc38369 |
| hc9217 |
| hc9222 |
| hc9454 |